


摘要
北极正在以惊人的速度变暖。虽然植物群落组成和物候的变化已经被广泛报道,但气候变化对生殖的影响仍然知之甚少。我们量化了格陵兰岛低北极和高北极地区九种苔原植物的花密度的多年代际变化。我们发现,随着时间的推移,花的密度发生了实质性的变化,但在不同的物种和地点,花的密度的时间趋势和驱动因素不同。低北极样地总花密度随时间增加,而高北极样地无方向性变化。在站点内部和站点之间,由于夏季温度、前一秋季温度和融雪时间的影响,物种之间的变化方向和速率不同。最后,所有物种在连续年份之间的花密度表现出强烈的权衡,这表明繁殖的有效成本。总的来说,我们的研究结果揭示了共同发生物种对共同气候驱动因素的敏感性和响应的区域和分类特异性差异,以及北极植物生殖投资的明确成本。因此,气候进一步变化的最终影响可能在物种之间和跨空间之间解耦,对植物物种动态、食物网结构和整体生态系统功能产生关键的连锁效应。
介绍
北极变暖的速度快于全球平均水平(IPCC 2021;vooosen 2021),对永久冻土、海洋和陆地冰表面动态、雪深和融雪模式以及生态系统功能有直接影响(Box等人,2019;Post et al. 2019)。在这种情况下,北极苔原生物群落提供了一个特殊的“自然实验室”来检验有关气候变化对生态群落影响的假设。尽管对北极植物物候变化及其性状响应气候变化进行了广泛的研究(Panchen and Gorelick 2017;prev
从植物相关分类群的角度来看,花是必不可少的资源。例如,许多北极昆虫依靠花作为食物,为植物提供授粉服务作为回报(Kevan 1972)。因此,花卉生产动态或植物群落组成的变化可以深刻影响当地生态网络和生态系统功能(Schmidt et al. 2016;Tiusanen et al. 2020)。考虑到北极地区最近和快速的环境变化(Robinson 2022),更好地了解决定花卉生产的机制对于量化和预测植物及其相关分类群的种群、群落和生态系统动态至关重要。
预测单个植物物种和群落如何应对气候变化是一个关键挑战。例如,在物种和地区之间,物候变化响应的速度和方向存在很大差异(Thackeray et al. 2016;Delgado et al. 2020;Roslin et al. 2021),包括冻土带生物群系(prev
除了非生物影响外,植物自身的年际资源分配策略也会影响长期的花卉生产(Obeso 2002;Høye et al. 2007a, b).在北极,大多数多年生物种在生长季节积累资源,在夏末或初秋产生花蕾。这种“芽前形成策略”相当于为下一个开花季节投入资源。早花物种通常受益于前一年形成的分化良好的花蕾,而晚花物种需要在生长季节芽成熟甚至完全形成芽(Körner 2003)。因此,某一年的花卉产量可能受到当年气候条件的影响,也可能受到前一年气候和用于繁殖的资源分配的影响。这种年与年之间的时间依赖性。,某一年与下一年开花之间的权衡可能会抵消当年开花的资源(Obeso 2002;Høye等,2007a;Semenchuk et al. 2013)。虽然变暖似乎导致苔原生态系统中群落水平开花季节的缩短(prev
使植物开花的时空格局更加复杂的是,北极不同地区的当前和预测气候条件存在很大差异(Abermann et al. 2017;prevac et al. 2017;Nabe-Nielsen et al. 2017;Bhatt et al. 2021)。由于在不同的气候条件下,环境的不同方面可能会受到限制,因此比较不同地点和不同条件下个体驱动因素对花卉生产的影响至关重要。
在这里,我们评估了在格陵兰岛两侧相距约1000公里的两个地点20年来花卉生产的时间变化和潜在驱动因素。为此,我们使用了9种典型北极苔原物种的花密度时间序列。考虑到气候变化和植被组成的空间异质性,我们检验了以下假设:
-
(H1)过去几十年的气候趋势在不同地点之间存在差异,表明不同气候变量在不同地点之间的变化率不同。
-
(H2)年花卉总产量随着时间的推移而增加,在变暖较快的地点增加更强。
-
(H3)不同气候变量对植物花产量的影响表现出不同的时间趋势。假设花卉生产模式与物候学报告的模式相似,我们预计一些物种将受益于较晚的融雪日期和温暖的前一年秋天,而其他物种将受益于较早的融雪和漫长而温暖的生长季节。
-
(H4)某一年的花卉产量与下一年的花卉产量呈权衡关系。在资源有限的情况下,我们预计今年在繁殖方面的投资增加将转化为下一年的投资减少。
材料与方法
花卉丰度和气候数据提取自格陵兰生态系统监测数据库(GEM, https://data.g-e-m.dk,提取于2022年)。
研究地点
为了比较不同气候条件和趋势的地区之间花卉生产的趋势和驱动因素,我们重点研究了格陵兰岛的两个地区,我们可以获得十年来的花卉丰度数据:位于低北极的Kobbefjord研究站(格陵兰岛西南部,64°08′N, 51°23′W),以及位于高北极的Zackenberg研究站(格陵兰岛东北部,74°29′N, 21°34′W,图1)。
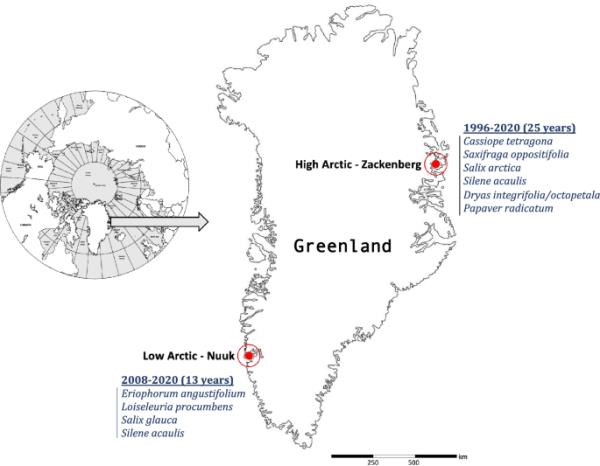
研究设计和数据范围。研究地点的地图,当地时间序列的长度和当地花丰度调查的物种列表
在低北极站点,2007-2020年期间,年平均温度为- 0.06°C。七月是最热的月份,平均气温为10.6°C(- 0.9至22.3°C);二月是最冷的月份,平均气温为- 8.6°C(- 30.2至9.2°C)。该地区多年冻土空间不连续,年平均降水量(雨雪总和)约838 mm。该地区陆地生态系统多样性丰富,包括低北极地区典型的植被类型:以桦树(Betula nana L.)、牛蒡(Vaccinium uliginosum L.)、黄杨(Empetrum sp.)、柳(Salix sp.)和四角仙桃(Cassiope tetragona L.)为主的沼泽、灌木林、草地和矮灌木荒原。D. Don (CAVM Team 2003;Walker et al. 2005)。在高北极地区,同一时期的年平均气温为- 8.6°C。七月是最热的月份,平均气温为6.8°C(- 1.9至19.6°C),而最冷的月份是三月,平均气温为- 20.9°C(- 38.4至2°C)。该地区多年冻土空间连续,年总降水量约260 mm。与低北极地区一样,可以发现各种苔原植被类型,从贫瘠的土地到沼泽和以矮灌木物种(如Cassiope tetragona (L.))为主的荒原。Don和Salix arctica Pall。
花朵丰度数据
开花数据由9种植物的年样地水平时间序列组成,其中生殖结构(如芽和花)的总数;统计每个样地在花季高峰期观察到的花的数量(研究区域和样地位置详见在线资源1)。每个样地用于记录9种监测植物中的特定植物物种,因此样地以下称为物种特异性样地。在监测开始时(即1996年为高北极地区,2008年为低北极地区),在积雪覆盖的局部梯度上建立了物种特异性样地,从而覆盖了物种生态位的全部范围。所有样地分别位于高北极和低北极的典型植被类型内。在高北极地区,在25年(1996-2020年)调查了6种植物,在低北极地区,在13年(2008-2020年)调查了4种植物(抽样设计见在线资源2)。所有监测项目包括的物种都是多年生植物:Saxifraga oppositifolia L.;棕枝露杉(L.)Desvaux;北极柳;青柳;桔梗;罂粟花;仙后座(L.)d .唐;青梅(L.)Jacq;以及杂交的综合森林树与八爪树(有关物种的更多详细信息,请参阅在线资源3)。
每周访问每个样地进行物候观察,并在开花高峰期记录生殖结构总数(Schmidt et al. 2019a);Raundrup et al. 2020)。地块被分为四个相同的部分(A-C),并使用绳索进一步细分地块以方便计数。所有的亚组都是在同一天统计的,但调查的确切日期因物种、样地和年份而异,这取决于当地的气候条件。被统计的生殖结构包括花、花蕾(柳属无性别)和衰老花(Schmidt et al. 2019a;Raundrup et al. 2020)。对柳属植物进行柳絮计数,每个样地分别记录雄性和雌性的总花数。由于芽是无性别的花,我们根据每个地块长期观察到的柳属花的平均性别比,独立地将其分为“雄性”或“雌性”。
由于样地大小差异很大(从1到300平方米不等),我们计算了每个物种的样地花密度,即样地内生殖结构的总数除以样地面积。对于立地水平的年开花密度分析(见下文SiteFD),我们将所有物种的年开花总数相加,并将其除以每个立地所有样地面积的总和。对于种级开花密度分析(见下文SpeciesFD),样地级花密度进行loge(x + 1)变换。
Environmental司机
我们提取了每个地点的当地气候数据。具体来说,我们使用了在站点水平记录的气温(°C)和累积降水(mm),因此每个站点的所有地块每年都有一个单一的值。这两个变量在Nuuk平均每30分钟记录一次,在Zackenberg每60分钟记录一次。因此,我们按日和月对每年的所有读数取平均值。我们计算了夏季(6 - 8月)和秋季(9 - 11月)的平均季节温度和降水,以及年平均温度和降水值,以估计每个站点的长期变化。低北极站点的时间跨度为2007-2020年,高北极站点的时间跨度为1995-2020年。
为了评估融雪日期,我们提取了在样地水平上记录的年度测量值,这些测量值是基于每个样地区域每周在野外进行1-3次的积雪目视估计。我们使用线性插值来预测一个地块(即所有部分的总和)达到50%积雪覆盖时的估计日期。请注意,在某些情况下,由于监测小组到达现场较晚,无法估计积雪覆盖50%的年份。
统计分析
我们的分析侧重于解决环境变化的四个印记:(i)每个站点气候条件的时间趋势,(ii)每个站点年总花密度的时间趋势(即每个站点内所有植物物种的总花密度),(iii)物种水平年花密度的时间趋势(每个站点内每个物种计算的花密度),以及(iv)前一年花密度的影响。以及温度和融雪日期对花密度的年际动态(每个站点内每个物种)的影响。
首先,为了检验高北极和低北极地区近期气候变化的差异,我们使用线性模型来量化年和季节平均温度和降水以及融雪日期的时间趋势。
其次,为了量化站点级别总花密度的时间变化方向和幅度(SiteFD, Eq. 1),我们将花密度建模为年份(Y,连续变量)的函数,并为每个站点拟合单独的线性模型。
(1)其中β1-2为估计参数,ε定义模型残差(假设为独立且正态分布)。在H1之后,我们预计花密度会随着时间的推移而显著增加,即β2 > 0。
第三,为了量化物种水平年花密度的时间变化方向和幅度(SpeciesFD, Eq. 2),我们将loge(x + 1)转换成物种之间双向相互作用的函数(S;分类变量)和年份(Y,连续变量)。我们为每个地点拟合了一个单独的线性混合模型。
(2)其中β1-4是估计参数,bplot是捕获plot影响的随机截距,ε是模型的残差(假设独立且正态分布)。注意,由于S是一个分类变量,其水平与每个站点的物种数量一样多,因此β2和β4实际上由多个估计参数组成。在H2之后,我们预计β4对每个物种都是显著的,即物种表现出显著不同的时间趋势。
第四,为了找出物种年花密度年际变化的驱动因素(SpeciesFD, Eq. 3),我们分别模拟了物种(S)与环境驱动因素(即当年夏季平均温度(ST)、一年融雪日(SM)和上一年秋季温度(PAT))之间的相互作用。我们进一步考虑了花密度的时间依赖性(TD),包括前一年的花密度(也是loge + 1转换)。Eq. 3中的不同术语旨在捕获我们旨在测试的多种气候效应,具体而言:ST用于捕获生长季节的温度效应;SM用于捕捉生长季节开始的效应;PAT用于捕捉上一生长期末资源积累的效应,TD用于捕捉上一年度繁殖成本的效应。由于花密度是在对数-对数尺度上建模的,因此连续几年的花密度相等,回归系数为1,而系数< 1意味着一年的高密度转化为下一年的低密度。因此,我们测试了TD与1之间的显著偏差(继Forchhammer et al. 2008之后)。从技术上讲,这是通过在模型中跨物种的TD上添加1的偏移量来实现的。在这方面,我们估计了1-β4而不是模型中的β4。因此,估计β4 > 1表明在时间t比时间t−1产生更多的花;β4 = 1表示每年产生的花数量相同,β4 < 1表示时间t产生的花比时间t−1少。换句话说,在我们的研究背景下,我们将β4解释为代表不同类型的时间依赖性(即补偿过多或补偿不足)。夏季温度(ST)、融雪日(SM)和往年秋季温度(PAT)均标准化为平均值0,标准差1。
(3)在Eq. 3中,对于Eq. 2, β1-4是与每个物种相关的估计参数的向量,ε定义模型的残差(假设为独立且正态分布)。另外,bplot和byear是随机截距,说明了plot在年内的交叉效应。,他们估计由实验设计的空间和时间结构引起的变化。值得注意的是,与其他参数相反,因为β4中的参数是针对1(而不是0)进行测试的,因此需要根据模型的这种特殊性计算与β4相关的p值,并使用“stats”包中的函数pt进行估计。
所有统计分析均在R (R Core Team 2022)中进行。线性混合效应模型使用“lme4”包中的函数lmer进行拟合(v.1.1-14, Bates et al. 2015)。
目录
摘要 介绍 材料与方法 结果 讨论 数据和材料的可用性 代码的可用性 参考文献 致谢 作者信息 道德声明 补充信息 搜索 导航 #####结果
气候变化
在低北极站点,12年的年、夏、秋平均气温变化趋势没有统计学意义(在线资源4)。然而,我们发现年平均降水量减少(在线资源4)。相反,在高北极站点,近25年的年、夏、秋气温和降水量增加(在线资源4)。低北极地区的年融雪日数没有变化,而高北极地区的年融雪日数没有变化。所有样地的积雪都倾向于更早融化(在Cassiope、Dryas、Papaver和Silene样地有统计支持的趋势)。
花卉总产量
高北极地区的平均年花密度远低于低北极地区(图2)。我们发现低北极地区的年花密度有所增加(β2 = 1.82花m−2年−1,CI[0.35,3.28], p = 0.02, r2 = 0.35),而高北极地区的年花密度没有变化(β2 = - 0.11, CI[- 0.27,0.05], p = 0.17,图2,Eq. 1)。
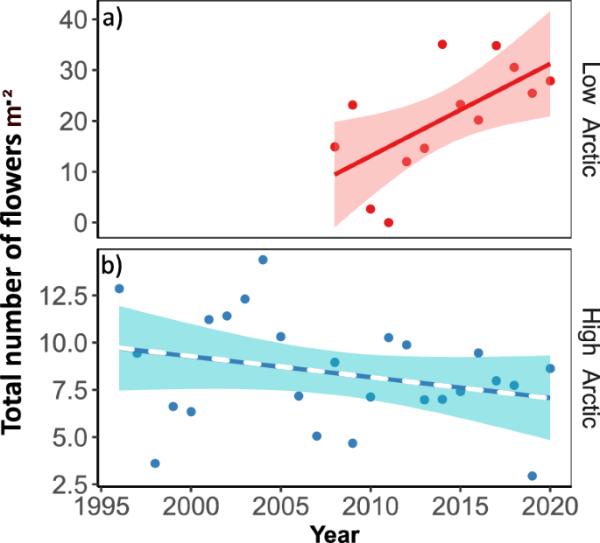
总花密度的时间趋势。图中显示的是a低北极地区和b高北极地区每个地点内所有物种的总数(即花的总数m - 2)。线条表示拟合回归,实线表示趋势显著,虚线表示影响不显著(根据Eq. 1);彩色带表示每个模型的估计置信区间。请注意两个图中y轴的不同尺度,高北极地区的平均花卉密度比低北极地区低得多
种级花卉生产
高北极地区:物种:年F = 2.84, p = 0.009;低北极地区:物种:年F = 8.42, p < 0.001, Eq. 2)。在低北极地区,我们观察到与Eriophorum和Salix-male相比,Salix-female、Silene和Loiseleuria的花密度呈增加的总体趋势,并且随着时间的推移没有明显的变化(图3)。相反,在高北极地区,所有物种的花密度都随着时间的推移而下降或保持稳定。Saxifraga和Papaver呈负趋势,而Cassiope, Dryas, Salix和Silene则没有明显变化,斜率估计接近于零(图3)。
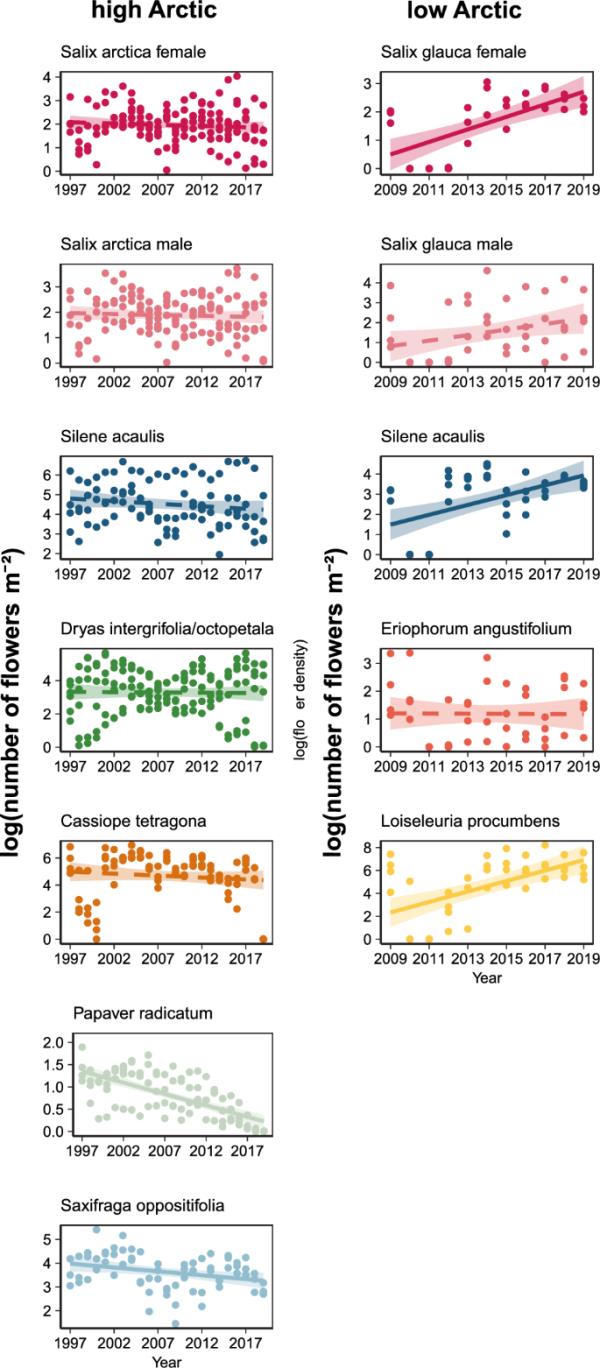
物种特有花密度的时间趋势。图中显示的是每个物种的(log + 1)花密度,分别为北极高和低地区。线条表示拟合回归,实线表示显著趋势,虚线表示非显著模型(基于Eq. 2);彩色带表示估计的置信区间。请注意,y轴的比例在面板之间是不同的
变革的驱动因素
在这两个样点上,前一年的花密度(TD)成为当年花密度的最重要驱动因素。这一效应对所有9个物种都是负的,即第t - 1年的花多意味着第t年的花少。然而,这种效应在物种之间的程度上存在显著差异(见表1中的物种:TD)。
气候驱动因素的影响高度依赖于物种和地点。只有三个物种经历了夏季温度(ST)的积极影响:低北极的Silene和Loiseleuria,以及高北极的Cassiope(见表1中的物种:ST)。高北极的Cassiope是唯一对先前秋季温度做出积极响应的物种(见表1中的物种:PAT)。最后,我们发现融雪日期对低北极的Loiseleuria,以及高北极的Salix-male和Cassiope有积极影响。在这些物种中,较晚的融雪反映在较高的花密度上(见表1中的物种:SM)。
在高北极地区,模型的固定效应包括不同的驱动因素(Eq. 3)解释了~ 10%的方差,而固定效应和随机效应联合解释了~ 95%的方差;低北极站点的对应值为~ 29%和~ 95%(表1)。
讨论
利用来自北极的独特的、长期的花朵丰度数据,我们的分析显示,随着时间的推移,花朵密度发生了显著的变化。然而,更具体的时间趋势和个别气候驱动因素的影响在地点和物种水平上都是不同的。总的来说,发现的模式与我们的先验假设是一致的。首先,我们发现在格陵兰岛两侧的地点之间,最近的气候和花卉生产趋势存在实质性差异。其次,地点水平上的花密度随着时间的推移而增加,但只在低北极地区如此。第三,我们发现了物种趋势的变化,不同的驱动因素对不同物种和不同地点的花密度起着重要的作用。最后,我们发现连续年份的花密度之间存在很强的权衡。下面,我们将依次讨论这些模式。
低北极地区和高北极地区近期气候变化的模式不同
当前的气候条件在北极不同地区之间差异很大,近期和预计变化条件的模式也是如此(Abermann et al. 2017;Kankaanpää et al. 2020;prevac et al. 2017;Schmidt et al. 2019b)。与这种区域差异相一致的是,我们的研究结果显示,格陵兰岛东北部的气候变化大于格陵兰岛西南部。
虽然低北极地区的年、夏季和秋季气温变化趋势不显著,但在研究期间,年平均气温从负向正变化(在线资源4)。这种气候条件的变化可能对植物生理以及土壤中水分和养分的有效性产生实质性的直接和间接影响。相比之下,在北极高纬度地区,年平均温度仍然是负的,尽管总体趋势是融雪日期提前和温度升高(即年、夏季和秋季平均温度)(在线资源4)。水在一年中大部分时间保持冻结,这一事实对养分和水分的可利用性以及植物代谢保持了强烈的限制。由于积雪减少,这些条件加上降水增加,使植物暴露在有害的冬季或春季冷空气温度下,从而对越冬组织和花蕾造成损害(Høye等人,2007年a;Inouye 2008;Semenchuk et al. 2013)。暴露在浅层积雪中的几种植物(包括Cassiope tetragona)在这种条件下会显著降低花的密度,而在八爪木(Dryas octopetala)中观察到的反应最强烈(Høye etal . 2007a;Semenchuk et al. 2013)。然而,在以雪的形式出现的强降水盛行的地方,在风没有将雪卷走的地方,一层厚厚的雪将在冬季结束时提供有效的保护覆盖物(Kelsey et al . 2021;Bokhorst et al . 2011)。
有限公司对比低北极和高北极花密度的时间趋势
随着温度的上升,我们预计北极高海拔地区的花密度趋势会更强——假设开花是由与物候相同的环境因素驱动的(Krab et al. 2018;Kelsey et al. 2021)。尽管如此,在过去的13年里,低北极地区的花的总密度几乎翻了一番,而高北极地区的趋势则不显著(或缓慢下降)(图2)。总花密度的解包装趋势揭示了植物物种和地点之间在方向和大小上的高度可变性。对于大多数低北极物种,开花密度随着时间的推移而增加,而在高北极地区,一些物种(如Papaver和Cassiope)呈现出强烈的下降趋势,而其他物种则没有明显的趋势(图3)。
花卉生产气候驱动因素的时空变化
在北极高海拔地区,我们没有发现明显的气候决定因素。晚花种Cassiope对气候变化的响应最大,其花密度随夏季温度、前秋季温度和融雪日期的增加而增加。这一发现与我们的先验预期一致,并强调了晚花物种在生长季节积累资源的必要性。在低北极样地,尽管Silene是晚花种,而Loiseleuria是早花种,但夏季温度对Silene和Loiseleuria都有积极影响。有趣的是,在北极高海拔地区,只有仙桃花和柳树雄花的丰度受到融雪时间的影响。然而,在低北极地区,早开花的路易色莱花似乎——正如预期的那样——受益于晚融雪。长时间放置的积雪可以保护预先形成的芽免受春季霜冻,并提供水分(Inouye 2008;Niittynen and Luoto 2018;Stewart et al. 2018)。随着时间的推移,我们没有发现环境驱动因素本身的显著趋势,即在监测Loiseleuria的地块中,融雪的天数。
年与年之间生殖投资的权衡与变异
我们发现在不同年份开花密度之间存在权衡的有力支持。具体来说,我们发现前一年的花卉丰度对今年的密度有一致的负面影响,与北极高海拔地区相比,低海拔地区的影响更大。这表明繁殖成本延迟,第1年花的对数-对数回归系数对第t - 1年密度的影响小于1(表1)。
由于北极花通常在前一年就已经形成(Körner 2003),我们可以预期,温暖的秋天会增加早花物种随后的花密度,因为较长的生长期应该允许它们在秋季获得额外的芽形成资源。然而,除了高北极地区的Cassiope外,所有物种似乎对这种影响都不敏感。对去年秋天的气温缺乏反应似乎令人惊讶,这可能是由于去年花卉生产的负面影响更大,而目前夏季条件的影响较小。
总体而言,北极环境条件具有强烈的时空变化特征。在目前的数据中,与年份和地块随机效应相关的高方差揭示了这一点(VarianceYear & VariancePlot: year,表1)。这种变化可能与影响湿度的微地形地块变化有关(Bannister et al. 2005;Høye等人,2007a)。这表明生活在该生态系统中的物种暴露于高度动态和异质性的环境中。因此,它们需要灵活和高效地利用资源,这一点可以从物种水平上的性状变异向更低温度的转变中看出(Siefert et al. 2015)。
花卉生产变化的意义
我们的分析显示,随着时间的推移,总花密度和物种特异性花密度都发生了实质性的变化。在低北极地区,我们观察到总体开花密度增加,而在低北极地区,开花密度下降的趋势是由沙弗拉加和Papaver驱动的。对于传粉者和其他与植物相关的分类类群来说,花朵数量的减少可能会增加获得花粉、花蜜或种子的竞争。在像北极这样资源有限的生态系统中,这种资源可用性的变化可以直接影响多样性和群落结构。例如,对植物来说,花卉产量的增加可以直接影响传粉媒介的竞争(hockking 1968;Tiusanen et al. 2020)。除此之外,植物相对物候学的变化可能会增加或减少竞争,这取决于变化如何影响更具吸引力和不太具有吸引力的植物物种之间的时间重叠(Høye et al. 2013;Tiusanen et al. 2020)。总体而言,正如在北极高纬度地区观察到的Saxifraga和Papaver所观察到的那样,未来花朵产量可能会减少,这有可能影响新个体的长期招募(Inouye 2008)和长距离传播。然而,由于许多苔原植物物种可能将有性生殖与无性繁殖和营养策略结合起来,因此很难将我们的结果与所有9种物种的种群动态直接联系起来(Wipf 2010)。需要进一步研究死亡率、定植、繁殖成功率和营养生长率,以将当前模式与生态系统尺度上的植物种群动态联系起来。我们目前的发现足以表明北极植物开花的主要变化,在物种和地点之间存在很大差异。
结论
本研究利用北极冻原9种植物的花丰度的长期时间序列,分析了气候变化和繁殖成本对花密度的时空影响。研究发现,在立地和种属水平上,过去和现在条件对花密度的时间趋势和影响存在差异。这种地点和物种水平响应的异质性突出了北极苔原植物群落中植被-气候相互作用的复杂性。这表明,当地群落以及共生物种可能对气候变化表现出截然不同的响应,因为它们对不同的特定气候驱动因素做出了响应。这些不对称模式可能影响植被动态,并导致其他营养水平的级联效应。每延伸一次,它们可能会在整个冻土带生态系统中产生反响。
下载原文档:https://link.springer.com/content/pdf/10.1007/s00300-023-03164-2.pdf