


摘要
气候变化对入侵风险的影响可能是消极的,也可能是积极的,这取决于物种。这些可以用物种分布模型进行预测,但预测结果会受到环境数据来源(气候数据源、全球环流模型GCM和共享社会经济路径SSP)的强烈影响。我们模拟了两种正在全球传播的马达加斯加爬行动物——大Phelsuma grandis和P. laticauda的分布。我们利用社会经济因素(例如,与港口的距离)和两个气候数据源,即地球和陆地表面区域高分辨率气候学(CHELSA)和Worldclim,来解释传播和建立的驱动因素。我们进一步量化了利用CHELSA和Worldclim数据对当前和未来条件的入侵风险模型的一致程度。大多数被确定为高度暴露于入侵风险的地区得到了一致的确定(例如加勒比和太平洋岛屿)。然而,预计的风险在各地有所不同。我们还发现,在当前条件下,数量入侵风险的差异也很显著(在适宜性评分上,阔叶树的差异为3%,而大叶树的差异高达14%)。尽管这两种物种的本地分布有很大的重叠,但到2070年,气候变化将导致相反的入侵风险响应(大叶茅减少,长叶茅增加)。总体而言,气候数据源对未来入侵风险预测的影响最大,其次是SSP。我们的研究结果强调,当前和未来入侵风险的评估对气候数据源敏感,特别是在岛屿上。我们强调在评估入侵风险时需要考虑多种气候。
介绍
外来入侵物种(IAS)正日益引起人们对其未来影响的关注,特别是由于其不断上升的经济成本和生态影响(Diagne et al. 2021)。气候变化对入侵风险的影响可能是积极的,也可能是消极的,这取决于物种(Bellard et al. 2013)。评估入侵风险及其如何受到气候变化的影响已成为制定积极保护行动的首要任务。
早期发现是防止入侵的关键决定因素,这表明迫切需要确定监测工作的优先领域。在这方面,保护生物学中广泛提倡的管理工具是物种分布模型(SDM;Gallien et al. 2012;Lanner et al. 2022)。该方法包括识别解释物种分布的环境因素和预测高环境适宜性区域。对于正在扩大的外来入侵生物,这种工具可以确定尚未被入侵但环境条件适宜且存在引进和传播因素的地区(例如海上交通)。IAS的分布可以通过环境和社会经济预测因素的组合来解释和预测(Bellard et al. 2016;Lanner et al. 2022)。环境预测因素(如气候和栖息地)可用于确定候选IAS可能进一步建立的领域,而社会经济预测因素(如港口和机场的邻近程度)代表传播和进入点的因素(Bertelsmeier和Courchamp 2014;Hulme 2021)。然后可以优先考虑入侵风险高的地区进行监测工作。
入侵风险可能会随着气候变化而变化,这可以使用sdm来评估,sdm包含了未来的气候预测(Bellard et al. 2013;Gillard et al. 2017)。这些预估根据所选择的情景而有所不同(例如,预估的全球社会经济变化情景,如共享社会经济路径(SSP),或温室气体排放情景,如代表性浓度路径(RCP))和全球环流模式(GCM,即方法论方面)。这些因素可以强烈影响sdm,并在气候适宜性评估中引起不确定性(Buisson et al. 2010)。最近,气候数据来源的选择(例如,CHELSA或Worldclim;菲克和希曼斯2017;Karger等人。2017)用于模型校准的θ已被确定为SDMs不确定性的主要来源(Baker等人,2016;Dubos et al. 2022a)。这些气候学的计算方法不同,导致最近一段时间气候变量的地理预测存在显著差异,这反过来又会极大地影响气候变化对物种分布影响的模式校准和预测。例如,基于Wordclim数据的预测表明,留尼汪岛的Phelsuma borbonica的气候适应性急剧下降,而在CHELSA上校准时几乎没有预测到任何影响(Dubos et al. 2022a)。与气候数据源相关的不确定性可能与GCM、预测者的选择(Dubos et al. 2022a)或建模技术(Stuart et al. 2022)的不确定性具有相似的量级。尽管如此,大多数旨在预测未来分布的SDM研究都使用来自单一来源的气候数据。据我们所知,还没有研究在入侵风险评估中考虑气候数据来源引起的不确定性。
在损害和管理方面,爬行动物是代价最高的入侵动物类群之一,在过去几十年里,估计每年的经济成本超过10亿美元(Diagne et. al. 2021)。爬行动物入侵的速度越来越快,以及相关的生态(例如,营养破坏)、进化/保护(例如,通过杂交或渗入)和卫生成本(例如,病原体传播)导致人们越来越关注某些物种(Reed和Kraus 2010;Sauteur et al. 2013;克劳斯2015;Vuillaume et al. 2015;Bellinati et al. 2022;Breuil et al. 2022)。然而,与无脊椎动物和植物物种相比,外来入侵爬行动物的研究仍然不足(例如,Bellard et al. 2013),需要填补气候变化如何影响爬行动物入侵风险的知识空白。
马达加斯加巨日壁虎(Phelsuma grandis Gray 1870)和金沙日壁虎(Phelsuma laticauda Boettger 1880)是两种分布在世界各地的马达加斯加爬行动物。大Phelsuma grandis是该属现存最大的物种之一,总长度可达30厘米(即大多数Phelsuma物种长度的两倍)。松林壁虎是一种中等大小的壁虎,体长可达13厘米;然而,该物种被认为是在法属波利尼西亚和留尼汪岛其他较小壁虎物种的积极竞争对手(Lund 2015;Deso et al. 2023),但也在其原生范围(Gehring et al. 2010)。由于它们的人为传播及其对土著社区造成的风险,这两种疾病都被认为是在其原生范围之外的外来入侵生物,包括马达加斯加中东部、毛里求斯、留尼汪岛、佛罗里达、法属波利尼西亚和夏威夷地区(Ota和Ineich, 2006年;Krysko and Borgia 2012;杜波2013;Buckland et al. 2014;Dubos et al. 2014;伦德2015;Fieldsend and Krysko 2019;Fieldsend等人。2020,2021c)。Phelsuma spp.(或其他爬行动物共享类似的栖息地,如变色蜥)的共存可能会通过竞争导致栖息地使用的变化(Harmon et al. 2007;Porcel等人,2021;Wright et al. 2021),这可能对更专门的本地物种有害。在毛里求斯,大叶黄杉的引入与四个地方性杉木种群的灭绝有关(Buckland et al. 2014)。众所周知,大壁虎、扁壁虎以及其他几种石斑虎(例如,高知壁虎)都会捕食体型较小的壁虎(Gehring et al. 2010;Buckland et al. 2014;Rakotozafy 2019),这表明对较小物种或类似大小物种的幼崽存在潜在的捕食风险。它们的引入也引起了人们对疾病和寄生虫传播给本地物种的风险的担忧(Dervin等人,2014;Barnett et al. 2018;Fieldsend and Krysko 2019;Fieldsend等。2021b;Unger et al. 2022),尽管迄今为止没有发现跨物种感染的证据(Goldberg and Bursey 2000;Leinwand et al. 2005)。大P. grandis和大P. laticauda的传播引起了人们对马达加斯加、毛里求斯和留尼汪岛本地(通常是地方性)动物群保护状况的更多关注(Dubos 2013;Buckland et al. 2014;Dubos et al. 2014)。它们与本地木杉物种的共存引起了人们对P. lineata、P. serraticauda、P. inexpectata、P. borbonica、P. cepediana、P. guimbeaui、P. ornata和P. rosagularis长期存在的关注(Andreone et al. 2003;格劳和旺斯2007;D 'Cruze et al. 2009;D 'Cruze and Kumar 2011;Blumgart et al. 2017;Porcel et al. 2021)。本文所考虑的这两种IAS普遍存在于城市化地区、观赏植物和果园以及原始雨林中,反映了巨大的生态位灵活性,这可能有助于解释成功的建立(Fieldsend等人,2021a)。这说明有必要描述它们的气候生态位,以便在全球范围内确定有入侵风险的潜在地区。
在此基础上,我们模拟了在当前气候条件和预测未来气候条件下大叶茅和大叶茅的分布,并预测了它们在全球范围内的入侵风险。为了确定气候数据源的选择是否会影响入侵风险评估,我们基于全球范围内两个主要气候数据源(即CHELSA和Worldclim)量化了当前入侵风险之间的一致程度。我们考虑了每个气候数据的多个不确定性来源,包括两个不同的气候情景(基于SSP情景),并对每个情景使用来自耦合模式比较项目6的多个环流模式(GCMs)。为了提供最可靠的保护指导方针,我们确定了两种气候数据源得出的预测之间一致的区域,并指出了优先监测的区域,以提高早期发现和预防潜在入侵的机会。
方法
大桫椤和桫椤均存在于多种生境类型中,包括原生林、高度退化的森林、果园和城市化生境(D’cruze et al. 2009;D 'Cruze and Kumar 2011;Dubos et al. 2014;Blumgart et al. 2017)。因此,我们假设栖息地变量不能很好地预测其环境,而气候变量可以更好地预测物种的分布(例如,Fieldsend等人。2021a)。我们的分析包括社会经济因素,如靠近道路、港口和机场,这些都是传播或潜在进入点的因素。
发生数据
我们从文献和机会观察中检索了本地和非本地范围(马达加斯加、毛里求斯、留尼汪岛、佛罗里达、法属波利尼西亚和夏威夷)的发生数据;格劳和旺斯2007;Raxworthy et al. 2007;Pearson and Raxworthy 2009;杜波2013;Buckland et al. 2014;Dubos et al. 2014;Fieldsend and Krysko 2019;Fieldsend等。2021a, 2021c;Porcel et al. 2021),所有这些数据都被纳入模型校准。结果表明:我们共获得338条独特的发生记录,而113条独特的发生记录。我们对数据进行了细化,以避免伪复制并减轻空间偏差,在环境变量的分辨率下,选择每像素出现一次(5角分,见下文)。这导致基于CHELSA的样本中有91个大斑拟虫存在点,其中50个在原生区域,41个在非原生区域(90个基于Worldclim;原生和非原生地区分别为49分和41分)。最终样本共有58个存在点,其中19个分布在原生地,39个分布在非原生地(59个基于Worldclim;原生和非原生地区分别为18分和41分)。
气候数据
我们使用了19个生物气候变量(描述可在https://www.worldclim.org/data/bioclim.html;另见Booth et al. 2014)以5角分(约10公里)分辨率从两个来源获得当前和未来(2070年)气候:CHELSA 1.2版(Karger et al. 2017)和Worldclim全球气候数据2.1版(Fick and Hijmans 2017)和1.4版,用于未来预测(与模式计算时CHELSA中可用的GCMs相匹配)。这些数据源使用不同的方法计算气候学,得到不同的地图(见图S1)。Worldclim基于除卫星数据外,以海拔和到海岸的距离作为预测因子的插值数据(Fick and Hijmans 2017),而CHELSA基于统计降尺度的温度和降水估计,其中包括地形因素(即风场、山谷显示、边界层高度;Karger et al. 2017)。我们决定包括所有19个生物气候变量,因为温度和降水都与物种的生物学有关,我们使用统计过程来选择最相关的变量(见下文)。对于每个气候数据源,我们为每组相互相关的变量选择一个预测变量,以避免共线性(Pearson’s r > 0.7;Dormann et al. 2013)使用虚拟物种R包的removeCollinearity函数(Leroy et al. 2016)。当平均值与极值共线时,我们选择了代表极端条件的变量(例如,给定时期内最温暖/最干燥的条件),因为这些变量更有可能导致死亡率和局部灭绝,并且与物种的建立有因果关系(Parmesan et al. 2000;Mazzotti et al. 2016;Maxwell et al. 2019)
对于未来的预测,我们使用了三个全球环流模式(GCMs);即BCC-CSM1-1、MIROC5和HadGEM2-AO)和两种温室气体排放情景(最乐观的RCP26和最悲观的RCP85),以考虑2070年可能的入侵风险。我们为两个气候数据源选择了相同的gcm和情景,以便能够量化与这些方面相关的变化(见下文“量化气候数据之间当前入侵风险的一致程度”小节)。
Socio-eco经济因素
我们使用到港口和机场的距离作为传播压力的引入因素和代理(Bellard et al. 2016)。我们从世界港口指数(http://k1.fpubli.cc/file/upload/202309/01/sgjdmmxu4a3)获得港口数据。从OpenFlights机场数据库(https://openflights.org/data.html,于2020年12月访问)获取机场数据。我们使用到主要道路和高速公路的距离作为潜在传播的指标,因为杉木物种可以被陆地车辆在短距离内意外运输(Deso 2001)。我们选择了道路规模最大的两类(高速公路和主要道路),并使用全球道路清单项目(GRIP4)数据集(Meijer et al. 2018)计算了与道路的距离。
分布模型
我们使用集合模型方法(四种建模技术)建模和预测物种分布。我们根据Valavi et al.(2021)选择了一组性能最好的建模技术。这些是随机森林下采样(RF下采样,即RF参数化以处理大量背景样本和少量存在记录);Prasad等人,2006),以及生物平台中三个表现最好的模型(Thuiller等人,2009):最近实现的MaxEnt,即MaxNet (Phillips 2017),广义增强回归模型(GBM,也称为增强回归树,BRT;Elith et al. 2008)和广义加性模型(GAM;Guisan et al. 2002)。RF下采样设置为运行1000个引导采样/树。
我们的数据集由仅存在的数据组成。因此,我们在从未检测到该物种的位置生成了伪缺席(Sillero et al. 2021)。我们首先生成五组不同的50,000个随机选择的伪缺席(或背景点)。我们的发生数据是从机会性观测中获取的,因此受到空间偏差的影响(例如,在人口稠密或可到达的地区附近更多的观测)。为了解释样本偏差,我们重新执行了基于不同伪缺失生成策略的校正的所有计算(需要校正和未校正的模型来可靠地测量样本偏差校正的效果;Dubos et al. 2022c;下面的细节)。在修正后的模型中,我们按照Phillips等人(2009)的做法,在存在点周围产生了五组伪缺席,以再现样本的空间偏差。我们使用由dismo R包(Hijmans 2012)生成的零地理模型(即到存在点的地理距离的地图),并将其用作伪缺席选择的概率权重。这种技术被认为适用于仍在扩展的IAS(即,不在平衡状态),因为它减少了合适但尚未入侵的区域(例如,Lanner et al. 2022)中伪缺席的产生。由于没有独立的数据可用于评估样本偏差校正的效果,我们使用了基于Schoener 's D重叠(Dubos et al. 2022c)的相对重叠指数(ROI)。ROI能够评估与模型运行之间的可变性相比,修正的影响是否可以忽略不计。它计算(1)未校正和校正预测之间的平均重叠(即校正的绝对效果),以及(2)每对模型重复之间的重叠(在每个伪缺失和交叉验证运行之间,分别针对每种建模技术,即模型随机性)。我们计算ROI如下:
其中为校正组模型运行之间的平均重叠部分,为未校正模型与校正模型运行之间的平均重叠部分。接近0的值表示校正的影响可以忽略不计(即样本偏差校正的影响与模型随机性的影响大小相同)。接近1的值表示校正效果为一周,模型随机性强。负值表明修正效应的量级低于模型随机性,因此不相关。如果未校正组和校正组之间的重叠小于运行之间的重叠(即ROI > 0;Dubos et al. 2022c)。
我们使用统计方法选择环境预测因子,将不相关的变量纳入我们对物种建立或传播的因果关系的假设中。对于每个气候数据源和物种,我们分别评估了每种建模技术以30种排列方式保留的每个变量的相对重要性(每个变量、数据源和物种总共= 120)。最终模型中包含的变量是那些相对重要性最高的变量。这些是使用肘标准在变量重要性的上铰链选择的(即每个变量的25%最佳表现模型),根据Harrell等人(1996)提出的“观察数/10预测因子”经验法则,为P. grandis设置最多9个变量,为P. laticauda选择5个变量(另见Guisan和Zimmermann 2000))。我们为当前分布计算了每个物种400个模型(4种建模技术× 5次伪缺失运行× 5次交叉验证运行× 2种样本偏差校正模式× 2个气候数据源),并利用未来气候数据计算了2400个预测(400个模型× 3个gcm × 2个ssp)。
模式e估值
通常建议使用空间划分来减少训练数据和测试数据之间的空间自相关性(即块交叉验证;Valavi et al. 2019)。在我们的例子中,发生数据是高度聚合的,这导致了块之间的严重不平衡。因此,我们对数据进行随机分区,80%的数据用于模型校准(训练),20%用于模型评估(测试)。该过程对每个物种、伪缺失数据集、校正方式和气候数据源重复了5次(交叉验证运行)。我们使用Boyce指数(Hirzel et al. 2006)来评估模型性能,该指数被认为是伪缺席数据的最佳评估指标(Leroy et al. 2018)。博伊斯指数值为1表示模型对存在点的预测较好,值为0表示模型的性能不优于随机。对于集成模型(即,跨建模技术、伪缺席运行和高性能模型的交叉验证运行的平均预测),我们丢弃了Boyce指数低于0.5的模型。
量化气候数据之间对当前入侵风险的一致程度
分别处理每个物种和样本偏差校正模式,我们比较了从CHELSA和Worldclim数据中获得的预测当前入侵风险。首先,通过计算集合图中所有像素适宜度之和的绝对差值,计算出每个物种CHELSA与Worldclim的差异指数;我们将这个差异除以CHELSA数据集所有像素的适宜性值的总和,以便将结果表示为Worldclim预测与CHELSA预测的绝对差异的百分比:
其中,PCHELSA和PWorldclim分别为CHELSA和Worldclim投影对像素j的适宜性评分。这种适宜性得分的差异表明,在整个预测区域内,两个预测之间的总体一致程度。适宜性分值差异大表明气候数据源的影响较大,无论是在总体适宜性分值方面还是在适宜环境表面方面。
其次,我们计算了一个考虑空间信息的替代指数,即空间重叠(Muscatello et al. 2021;Petford and Alexander 2021;Dubos et al. 2022a)。我们计算了基于考虑的两个气候数据集的预测之间当前入侵风险预测之间的Schoener’s D重叠。值为1表示产生的两个预估在空间上完全匹配(即没有气候数据源的影响),值为0表示完全不匹配。我们使用ENMTools R包计算了CHELSA和Worldclim预测之间的Schoener 's D重叠(Warren et al. 2010)。Schoener’s D的计算公式如下:
其中pxi和pyi是网格单元i中未校正x和校正y预测的归一化适用性分数,对于每个物种,交叉验证运行和伪缺席单独运行。
确定监测的优先事项
为了确定入侵风险最高的地区,我们根据前面步骤中每种物种的入侵风险量化对国家和岛屿进行了排名。我们使用了从Global ADMinistrative area (GADM v4.0.4;https://gadm.org/data.html)将预测的入侵风险与相应的国家、岛屿或群岛(所有具有ISO国家代码的领土,例如马达加斯加、留尼汪岛、科摩罗)联系起来。然后我们计算了每个地区的平均入侵风险。为此,我们提取了集合模型的预测值,并在区域的所有像素上取平均值。这种方法可能会淡化只有小区域有风险的大国的风险,但对我们的研究物种是有用的,这些物种主要在小岛屿上发现。由于我们对哪个气候数据源最适合生态建模没有先验预期,我们基于CHELSA和Worldclim之间预测的平均值进行排名。为了考虑不确定性,我们通过减去其标准差(mean - sd)来惩罚平均预测,遵循Kujala等人(2013)开发的方法,适用于单一物种(Dubos等人,2022a)。这使我们能够优先考虑在气候数据源和模型复制中最一致地确定为高入侵风险的地区。我们将这些受惩罚的平均预测称为共识入侵风险。为了比较,我们还提供了从两个单独的气候数据源获得的排名(表格可在辅助信息中找到)。
预测未来气候变化的影响
我们将模型的预测值与未来的气候数据相结合。对于每个物种、气候数据、GCM和情景,我们分别用上述两种互补方法量化了当前和未来预测入侵风险之间的差异,即总(未受惩罚的)适宜性得分和空间重叠的差异(Dubos et al. 2022a)。我们计算了入侵风险的两个变化指数(我们进一步将这些指数称为物种范围变化SRC,继Buisson等人2010年和Baker等人2016年之后)。我们首先计算了未来和当前预测的总适宜性得分之间的差异,并显示了相对于当前适宜性得分的增加/减少比例。其次,我们使用Schoener 's D重叠来量化空间适宜性变化,以解释空间信息(例如,即使在总体适宜性没有变化的情况下,也允许我们识别分布变化)。我们验证了模型在使用夹紧口罩和检查预测器响应的形状来预测新的(未来的)数据方面得到了充分的信息。
量化未来预估中与气候数据有关的不确定性
我们量化了与气候情景、gcm和气候数据源相关的SRC(总得分和Schoener’s D的差异)的不确定性。对于总得分的差异,我们使用线性模型(LM,假设高斯误差)量化气候数据模式解释的偏差比例,以SRC作为响应变量,并将上述不确定性来源作为解释变量,遵循Baker等人(2016)。然后,我们评估了由每个不确定性来源f解释的偏差比例,如下所示:
式中,Pf =被因子f解释的偏差比例,D1 =全模型偏差,Df =全模型偏差减去因子f, D0 =零模型偏差。
我们使用β -回归GLM代替LM对Schoener 's D重叠重复了这一分析,因为重叠度量的范围连续在0到1之间(glmmTMB R包;Brooks et al. 2019)。
目录
摘要 介绍 方法 结果 讨论 数据可用性 参考文献 致谢 作者信息 道德声明 补充信息 搜索 导航 #####结果
物种分布模型
根据气候数据源的不同,我们选择了6到7个变量作为大P., 5个变量作为大P.(表1;图S3-S10)。这两种物种的当前分布最好由社会经济变量“到港口的距离”和/或“到机场的距离”(表1)和与温度变化率(日差bio2和温度季节性bio4)或最低温度(bio6)相关的气候变量来解释。最暖季降水(bio18)对大叶藻也很重要。对于这个物种,我们放弃了“到道路的距离”,因为这个预测产生了虚假的结果,这与我们的生物学假设(即,100公里距离后入侵风险增加)不符,并且会降低我们模型的可转移性。两个物种均分布在港口附近(约250公里以内)和机场附近(约250公里以内)。在100公里范围内),在温度变率低、最低气温高(最冷月份平均> 10°C)的地区,以及在大叶蝉的情况下,避开干旱地区(夏季降水> 250 mm);图S5-S6、S9-S10)。
模型通常在预测已知存在方面做得很好(大多数Boyce指数> 0.5),与基于chelsa的预测相比,基于worldclimm的预测具有更高的Boyce指数(表2;图S11-S12)。抽样偏差校正的影响比模型随机性更重要(CHELSA的ROI = 0.07, Worldclim的ROI = 0.09)。在每种模式(物种、气候数据源和样本偏差校正)的100个模型中,我们丢弃了5到25个表现不佳的模型;表2)。
当前入侵风险
我们在世界各地的多个地区发现了重要的入侵风险,主要是在热带岛屿(图1,2)。在这两个物种中,我们发现印度洋岛屿(例如科摩罗,马约特岛),太平洋(例如纽埃,新喀里多尼亚),加勒比地区(特别是在大安的列斯群岛和巴哈马群岛),中非两海岸(安哥拉,刚果,坦桑尼亚,莫桑比克)和印度太平洋地区(菲律宾,越南,新几内亚)的入侵风险最高。佛得角和巴西海岸也有适合这两个物种生存的条件。我们发现在小安的列斯群岛、瓦努阿图和夏威夷群岛,这两个物种之间的入侵风险不同,在这些地区,P. laticauda的入侵风险更大(图3)。
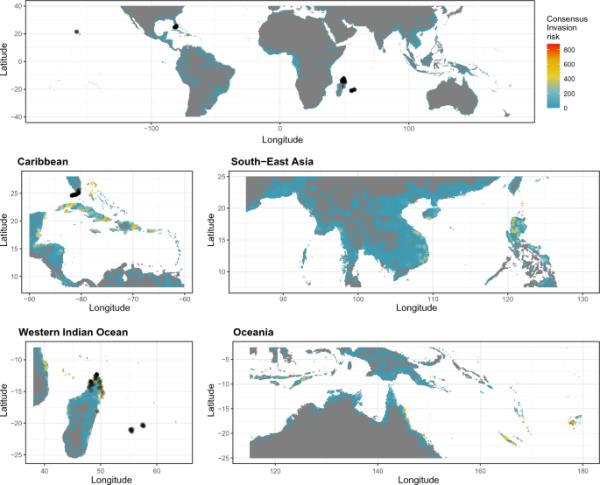
目前的共识是大Phelsuma的全球入侵风险。从所有模拟(包括基于CHELSA和Worldclim气候数据的模型)的平均预测中得出一致的入侵风险,减去标准偏差以考虑不确定性。封闭的黑色圆圈表示存在点。气候数据源特定地图可在辅助信息中找到(图S13-S16)
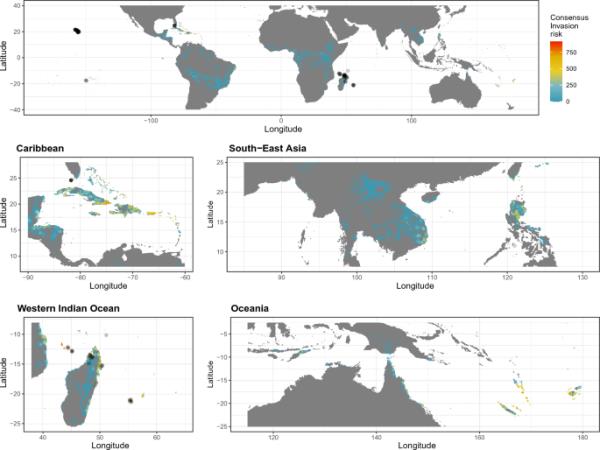
目前的共识是松柏的全球入侵风险。从所有模拟(包括基于CHELSA和Worldclim气候数据的模型)的平均预测中得出一致的入侵风险,减去标准偏差以考虑不确定性。封闭的黑色圆圈表示存在点。气候数据源特定地图可在辅助信息中找到(图S17-S20)。
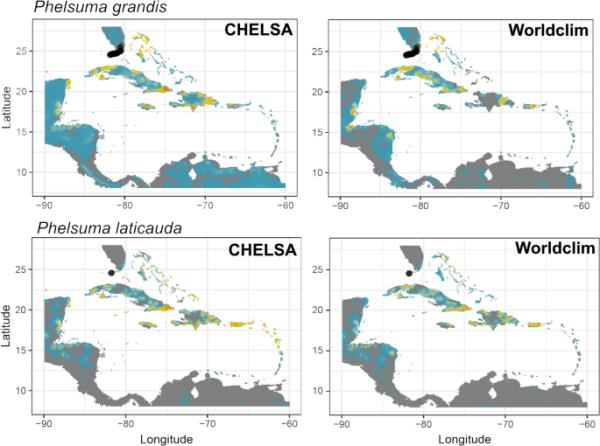
基于两个气候数据来源(左:CHELSA;右图:世界气候)在加勒比地区。预测值在模型重复中取平均值(每个物种和气候数据源的n = 100),并受到不确定性的影响(重复间的标准变异)。红色代表高确定性的高入侵风险,蓝色代表中等入侵风险,灰色代表不确定性高于入侵风险的区域
当使用不同的气候源进行校准时,当前入侵风险的预测在当地有所不同(见图3中的加勒比地区的例子;单个集成模型可在支持信息中获得,图S13-S20)。我们发现,两种预测结果在总适宜性得分和空间重叠方面存在显著差异(表3)。气候数据源影响了每一领土的入侵风险排名,有时差异很大(例如,大蠊在圣赫勒拿岛、阿森松岛和特里斯坦达库尼亚的排名差异为36;P. laticauda的圣马丁等级差30;表S1,表S2)。
未来的入侵风险
我们预测在大多数情况下,到2070年入侵风险会降低(图4;S19-22)。我们发现基于不同气候数据的预测之间存在重要差异,基于worldclims的预测总体上具有更高的SRC(图4)。对于大蠊,我们预测所有情况下的总入侵风险降低,范围在8.6 - 16.1%(总分)之间,空间变化范围在9.9 - 19.4%之间(1-Schoener’s D重叠),具体取决于情景和气候数据。不同气候数据源对柽柳的气候变化效应预估差异较大,总体入侵风险在- 4.7%(下降)~ + 18.8%(增加)之间,空间变化幅度在11.7% ~ 20.8%之间。在整个原生和入侵范围内,仅对Worldclim(图S25, S26),夹持面罩表明了一个变量的新条件。这些新条件似乎主要是由最高温度驱动的(Bio5;图S27)。考虑到我们的预测因子与适宜性之间的驼峰状关系(图S5、S9、S10),明确地确定了气候条件的合适窗口,由于外推而导致预测不确定的风险很小。
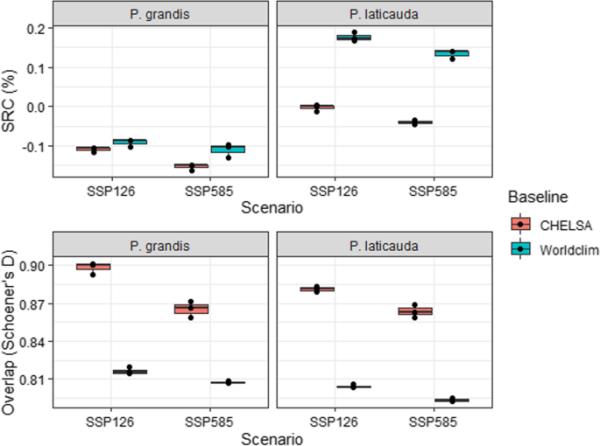
气候数据源对入侵大叶茅和长叶茅物种范围变化(SRC,当前适宜性与未来适宜性之间的总体差异,此处以相对于当前适宜性的比例表示)和Schoener’s D重叠(当前与未来预测之间共同空间信息的百分比)的影响对模型进行样本偏差校正。为每个气候数据源、情景(SSP)和全球环流模式(GCMs,用黑点表示)分别计算了指数。方框代表第25和75个百分位,柱状图代表中位数
在大多数情况下,气候数据来源是未来入侵风险预测中最重要的不确定性驱动因素(图5)。就总分(SRC)而言,气候数据来源解释的不确定性与柽柳情景解释的不确定性一样多。此外,气候数据源是影响两种物种空间重叠不确定性的最大驱动因素。
.jpg)
气候数据模式(情景、全球环流模式和气候数据源)在预估气候变化效应(重叠和适宜性评分)中对a大林柏和b小林柏的影响的偏差比例用Schoener 's D计算当前和未来预测之间的重叠;适宜性分数是当前和未来预测的总分之差
讨论
我们使用最近提倡的方法模拟了两种入侵的外来爬行动物物种的当前和未来分布,考虑了社会经济因素和广泛的气候数据。我们确定了几个入侵风险较高的地区,这些发现对气候数据的选择是强有力的。我们建议将这些地区视为监测工作和监测的重点地区,但也必须考虑由单一气候数据确定的有风险的地区。我们发现了与气候数据来源有关的重要差异。总体而言,我们预测气候变化将降低大叶茅的入侵风险,略微增加叶茅的入侵风险。
传播和建立的驱动因素
这两个物种的传播都是由海上和/或空中运输驱动的。靠近港口和机场的重要性可能是由于我们研究物种的孤立性(大多数岛屿都有港口和机场)。然而,使用我们的伪缺席采样策略(更集中在存在点附近),这些变量的这种影响的检测表明,物种在港口和机场附近的存在比偶然预期的要多。模式确定了距离较远(分别为250公里和100公里)的港口和机场附近的影响,这可能是由位于大陆地区(马达加斯加和佛罗里达)的发生点驱动的。这也许可以解释小岛屿(如奥胡岛)几乎持续的高入侵风险。然而,我们仍然确定了宽度小于100公里的岛屿(如夏威夷和留尼汪岛)内入侵风险的梯度,这表明有可能优先考虑小岛屿的特定区域。在酒店和植物苗圃等人为结构上,均可发现大叶蝉和大叶蝉。奥林匹克广播服务公司。、伊内希、乔尔、克罗蒂尼;Gehring et al. 2010),以及香蕉和椰子等观赏植物和种植园作物(Gill et al. 2001;D 'Cruze et al. 2009;Porcel等人,2021)。因此,个体可能经常通过集装箱运输,并意外地引入运输(Fritts 1987;Dubos et al. 2014;Khoury等人。2021)。在为宠物贸易进口它们的地区(例如,在佛罗里达州;Andreone et al. 2012;Fieldsend and Krysko 2019)。我们没有将宠物贸易本身纳入我们的分析,因为宠物贸易的影响主要是由于运输(通过港口和机场的集装箱意外释放,就像马达加斯加的Toamasina一样;Dubos et al. 2014)。因此,宠物贸易的影响已经考虑到我们的模型与接近港口和机场的预测。此外,与商业化后的宠物贸易相关的引进主要是有意的,这只在佛罗里达州被观察到。因此,添加宠物贸易数据可能会导致我们的结果出现偏差。在日和年尺度上,这两个物种都能够在温度变化较小的地区定居。它们的原生地位于马达加斯加北部,靠近海岸,那里的气候主要是赤道气候(Peel et al. 2007),年和日热变化很小。它们与低热变异性的亲和力可能与温度波动对活动和繁殖的强烈影响有关(Georges 2013;Noble et al. 2018, Choeur et al. 2022)。我们研究的物种全年都很活跃,这可能解释了它们与低温季节性的亲和力。它们全年的活动意味着需要持续的食物供应。低季节性可能有助于维持果实/花蜜的生产和昆虫活动,这些都是大叶茅和叶茅的食物来源(Dervin et al. 2013;Dubos et al. 2020;horaau et al. 2021)。在日尺度和年尺度上与低变异性的联系可能是由于温度依赖的性别决定,这是包括Phelsuma spp在内的爬行动物的共同特征(Gamble 2010;Cornejo-Páramo et al. 2020)。大斑拟虫和扁斑拟虫都把卵产在基质表面,使它们暴露在每天温度波动的环境中。在我们的情况下,白天和全年相对恒定的温度可能有助于平衡性别比例和维持人口动态(Georges 2013)。
我们发现,我们的研究物种无法在最低温度(< 10°C)的地区建立,可能是因为寒冷降低了变温动物的活动,从而降低了它们的生存能力。这对应于热带爬行动物中常见的热耐受性的下界(Sunday et al. 2011)。最低温度也可能影响孵化时间和性别决定(Georges 2013;Roesch et al. 2021)。延长的孵化期可能会增加孵化失败和蛋被捕食的可能性。较低的筑巢成功率和不平衡的性别比例可能会破坏种群动态,阻止它们在寒冷地区的生存。考虑到与马达加斯加北部相比,佛罗里达州冬季偶尔出现的低温,这两个物种的出现可能令人惊讶。最近对大叶茅气候生态位的评估显示,大叶茅原产地的气候与佛罗里达被入侵地区的气候存在重要差异(Fieldsend et al. 2021a)。这表明热塑性或适应新环境的潜力很大(Card et al. 2018;Lapwong et al. 2021)的基础是物种的本地分布受到生物相互作用(捕食和竞争;例如,与P. kochi的竞争;Fieldsend等人。2021a)。这与对frenatus半趾蜱和Anolis sagrei的研究结果一致,它们能够在比其原生范围更冷的地区传播(Angetter等人,2011;Lapwong et al. 2021)。高遗传多样性往往有助于入侵的成功(Angetter et al. 2011),而多种本地来源可能会增强入侵的成功(Fieldsend et al. 2021c),佛罗里达的P. grandis就是如此。进一步的研究可能会评估这两个物种在其入侵范围内的遗传多样性水平,以更好地了解物种在新环境中生存的能力。
这两种壁虎物种都没有在干旱季节的地区定居,可能是因为低降水限制了初级和次级生产,从而限制了食物供应(Dubos et al. 2019)。长时间的干旱期与爬行动物的身体状况下降、死亡率增加和局部灭绝有关(Maxwell et al. 2019),这些条件可能会阻碍我们研究物种的建立。
当前入侵风险
在CHELSA和基于worldclims的预测中,预测的高入侵风险地区是一致的,但在地方上存在重要差异。它们大多位于加勒比岛屿、西印度洋岛屿、东南亚和东大洋洲。外来木栓类的潜在入侵可能会使当地动物群面临新的竞争对手或捕食者。大叶茅和大叶茅在生境利用方面都具有高度的灵活性(D’cruze and Kumar 2011;Dubos et al. 2014),这引起了对本地共生物种以及生活在自然森林栖息地的物种的关注。面临危险的物种包括本地的石斑鱼物种(如东部海港城镇Toamasina的P. lineata种群减少所表明的),或任何其他具有类似栖息地利用的日间树栖爬行动物(例如,栖木高度、底物;Augros等人,2018;Wright et al. 2021),例如来自海地的极度濒危的棕色红腹变色蜥Anolis koopmani,来自海地和多米尼加共和国的濒临灭绝的黑喉黑斑变色蜥Anolis armouri,或来自古巴的极度濒危的Finca Ceres变色蜥Anolis juangundlachi。鉴于我们的研究物种所占据的栖息地类型范围广泛,还应关注所有分布与危险地区相匹配的较小物种(例如,来自新喀里多尼亚的Bavaya spp.或Eurodactylodes spp.)的保护问题。汤加极度濒危的“尤亚森林壁虎”Lepidodactylus euaensis尤其值得关注,因为它的保护状况和对这个岛屿的极高入侵风险。P. grandis和P. laticauda都是白天活动的,但由于人造光,它们也可以在夜间活动(Dubos et al. 2020;Baxter-Gilbert et al. 2021),强调了与生活在人类建筑附近的夜行物种(如极度濒危的巴巴多斯叶趾壁虎Phyllodactylus pulcher)竞争的风险(Williams et al. 2016)。入侵的杉木物种对本土动物的潜在影响可以通过潜在的可塑性来缓解,这可能会促进小气候和/或栖息地的划分(Noble et al. 2011;Porcel等人,2021;Ryan and Gunderson 2021)。未来的研究应进一步调查大叶茅、大叶茅及其同域物种在空间、时间或环境方面的潜在变化,以更好地了解哪些物种面临更大的风险。
气候数据源的选择
我们使用不同的气候数据源确定了预测入侵风险之间的局部差异。差异可能是由不同变量的选择驱动的(例如,用CHELSA数据校准的模型选择了“每日温度范围”,而没有用Worldclim来校准P. grandis);然而,最近的一项研究表明,即使选择了相同的预测因子,差异也会持续存在(Dubos et al. 2022a;另见jimsamnez - valverde et al. 2021)。这种不匹配可以用计算气候学的方法来更好地解释。Worldclim是由插值数据构建的,除了卫星数据外,还有海拔和到海岸的距离作为预测指标(Fick and Hijmans 2017)。CHELSA对温度进行了统计降尺度,降水估算纳入了地形因素(即风场、山谷暴露、边界层高度;Karger等人,2017)。在地形异质性较强的地区(如大洋岛屿),这种差异可能会加剧(例如,Lannuzel et al. 2021)。时间覆盖度的差异可能是另一个不匹配的来源,Worldclim代表1960-1990年期间的条件,而CHELSA计算的是1979-2013年。由于我们不知道哪种气候数据源对预测入侵风险最有用,我们建议,旨在预测当前和未来入侵风险的研究应考虑多种气候数据源,并量化与这些数据源相关的不确定性。
未来的入侵风险
总体而言(即考虑到SRC),根据Worldclim和CHELSA,我们预测未来的气候变化将降低大斑拟虫的入侵风险,正如入侵爬行动物通常发现的那样(Bellard et al. 2013;但参见Piquet et al. 2021)。请注意,在缺乏生物安全措施的情况下,尽管气候影响,入侵的可能性很大。另一方面,根据Worldclim的预测,柽柳的入侵风险会增加,而根据CHELSA的预测则不会增加。对于这两个物种,基于世界气候的预测比基于chelsa的预测更倾向于预测更高的风险。与Worldclim的空间不匹配(重叠)也更大。基于CHELSA和Worldclim的未来预测之间的差异与两个极端情景(SSP126和SSP585;这表明,纳入多个气候数据集与纳入排放情景同样重要。
爬行动物可能会改变其物候以响应环境变化(Kearney等人,2009),这已经在Phelsuma spp中观察到(Dubos等人,2020;Baxter-Gilbert et al. 2021)。对气候变化的行为反应——以及物候变化——可能与地理反应相互作用(Kearney et al. 2010)。为了充分了解入侵爬行动物对气候变化的反应,并采取积极的行动,还需要进一步的研究。
对采用5角分分辨率
海洋岛屿的特点是地形高度不均匀,在小区域内气候条件有很大的梯度。在全球范围内,高分辨率(例如30角秒)对计算能力和数据存储的要求非常高,这就是我们选择5角分分辨率的原因。我们的预测不能很好地代表这种异质性,我们不能在精细尺度上确定合适的区域。这可能导致高估了我们研究物种的气候生态位宽度。然而,我们的预测显示了热带小岛(如科摩罗、马约特岛和巴哈马群岛的小岛)之间的差异,这支持了我们确定的最符合物种气候生态位的地区。预估还显示了大于1个像元(> 10公里;例如,留尼汪岛和夏威夷岛;图S13, S14, S17, S18),表明在大多数情况下成功地排除了高海拔较冷气候。我们的目标是确定入侵风险高的地区和岛屿,以加强入境点的监测工作,而不是预测可能被占领的局部地区,我们相信我们的结果与全球范围内的优先级相关。
建模非均衡入侵风险
入侵物种的生态位模型是错误的,因为它们违反了平衡假设(Hui 2022)。然而,如果经过适当的校准,它们是有用的,因为它们可以根据对入侵物种分布的现有知识,提供适合入侵物种生存的地区的指示。为了使SDM预测成为入侵风险的有用指标,该协议必须考虑到缺乏均衡和其他偏差。有两种可能的方法来处理这个问题。第一种方法包括排除非本地事件(Barbet-Massin et al. 2018)。这一点已经在p.g grandis中进行了测试,并且基于没有本地发生点的校准模型的预测忽略了一些被占用的区域和合适的气候条件(Fieldsend et al. 2021a)。第二种方法是解决导致建模协议缺乏平衡的过程(Hui 2022)。我们从两个方法方面采用了第二种方法。我们添加了与引入和传播风险相关的具有明确假设的变量(例如Lanner et al. 2022)。通过添加这些变量(这里我们使用到港口和机场的距离),该模型不太可能在气候变量中寻找对物种范围的非本地限制的解释。同样,抽样偏差可能导致与非平衡违反类似的影响:如果不考虑偏差,那么模型将在气候变量中寻找解释,这将导致误导性的结论。我们在这里使用了一种样本偏差校正技术,该技术将伪缺席点的生成集中在存在点周围。这种方法减少了在未到达地区忽视合适环境的可能性。
有限公司ncluding讲话
直到最近,SDM研究才考虑到气候数据的来源(Baker et al. 2016;莫拉莱斯-巴贝罗和维加-Álvarez 2019;Datta et al. 2020;Ocon 2020;Dubos et al. 2022b, a;Stewart et al. 2022)。据我们所知,这项研究是第一个在入侵风险评估中考虑多种气候数据来源的研究。我们强调了环境适宜性量化的空间差异,这可能导致风险区域的遗漏。进一步的研究应该在更广泛的分类尺度和不同的景观(特别是较小的海洋岛屿与大陆)上评估入侵风险对气候数据的敏感性。
如果及早发现,IAS的经济成本很低,但如果不及时发现,则会由于造成的损害和管理努力的增加而迅速上升(Renault et al. 2021)。就全球年度经济成本而言,爬行动物是第二大入侵脊椎动物类别(Diagne et al. 2021)。因此,促进有效的生物安全措施以确保早期发现似乎在很大程度上在经济上是可行的(Cuthbert et al. 2022;Dubos et al. 2023)并培养公众意识以减少故意释放(Perry and Farmer 2011)。考虑到生态和经济方面的利害关系,应该考虑在根据单一气候数据确定为具有入侵高风险的地区实施监测规划。然而,监测工作应该优先放在高度确定的高入侵风险的地方,即基于考虑多种气候数据源的预测。
下载原文档:https://link.springer.com/content/pdf/10.1007/s10530-023-03082-8.pdf